Lymphocytes are the major player in adaptive immunity, or immunity that responds specifically to a pathogen. These lymphocytes have two broad categories: T lymphocytes and B lymphocytes.
T Lymphocytes
T lymphocytes mature in the thymus, hence the name “T” lymphocyte. After leaving the thymus, these lymphocytes can be found in the periarteriolar lymphoid sheaths of the spleen and in the paracortical regions of lymph nodes.
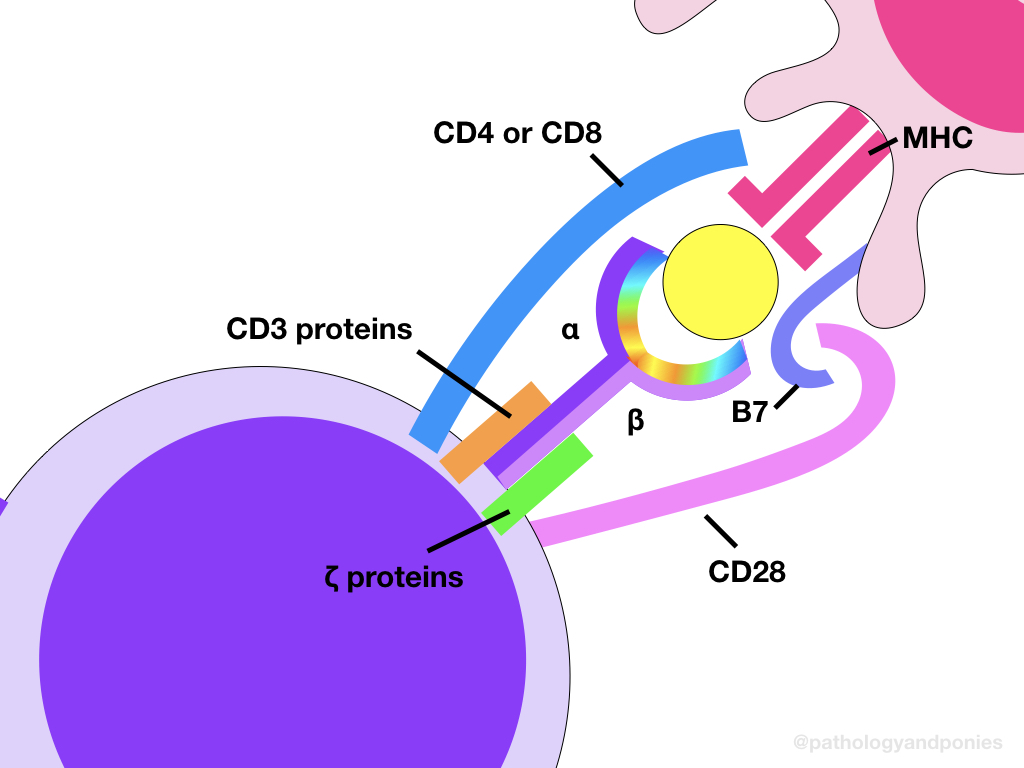
There are two main types of T lymphocytes, determined by their accessory CD molecule. Helper T lymphocytes express CD4, while cytotoxic T lymphocytes express CD8. These accessory molecules are attached to the T cell receptor (TCR) complex, which is the major activating receptor for T cells.
The TCR
The T cell receptor is the major antigen-binding receptor on T cells, allowing for the cell to be activated against a particular pathogen. Each TCR has a certain pathogen specificity, meaning that only TCRs with the correct genetic makeup to match a particular antigen will bind to that antigen, and only T lymphocytes expressing that specific TCR will be activated. The specificity of each TCR binding site is determined during maturation in the thymus, where somatic rearrangements produce variation in the binding site. This is discussed further in Immunologic Tolerance.
The basic TCR is made up of two linked heterodimers, which are either αβ or ɣδ. The relative proportion of αβ-TCR T lymphocytes versus ɣδ-TCR T lymphocytes varies between species. For example, ruminants and pigs have a higher proportion of ɣδ-TCR T lymphocytes. It has been suggested that having a higher proportion of ɣδ-TCRs means these species are more likely to form intestinal granulomas, because the method of activation of αβ and ɣδ-TCRs is different.
αβ-TCRs can only recognize antigens processed by cells and presented on MHC molecules. So, αβ-TCR T lymphocytes will only respond when activated by an antigen-presenting cell or a stressed cell expressing viral or neoplastic proteins. Conversely, ɣδ-TCRs can recognize antigen that is not presented by MHC. In most species, these cells are primarily found at epithelial surfaces, and may help provide an early cell-mediate immune response if epithelial integrity is lost. This is the proposed mechanism by which granulomas are more likely to form in ruminants and pigs, as these ɣδ-TCRs at the epithelial surface will induce a rapid granulomatous inflammation in response to pathogens like mycobacteria. However, the role of these ɣδ-TCRs is still somewhat controversial.
Attached to the main TCR heterodimer is a molecule called CD3 and two zeta (ζ) chains. These components are invariant, and aid in signalling when an antigen is bound to the TCR. Finally, the aforementioned accessory molecules, either CD4 or CD8, are also linked to the TCR complex. These molecules bind to a specific type of MHC molecule, giving the T lymphocyte either a cytotoxic or helper function.
A brief note on NK-T lymphocytes:
Besides αβ-TCRs and ɣδ-TCRs, there is actually a third type of T lymphocyte TCR that exists. These are NK-T lymphocytes, which express molecules similar to natural killer cells, but also express some TCRs. The exact function of these cells is unknown.
The Second Signal
Although not a part of the TCR complex, there is a second critical component to T lymphocyte activation. This is CD28, another protein expressed by T lymphocytes. When a T lymphocyte binds to another cell via the TCR, CD28 binds to a B7 protein, either CD80 or CD86, which is expressed on the target cell. This binding acts as a second signal that is required for T lymphocyte activation. This requirement is one of the ways that autoimmune disease is limited. This is discussed further in Immunologic Tolerance.
Helper T Cells (CD4⁺)
T lymphocytes with CD4 only bind to antigen presented on MHCII molecules. There are actually three types of helper T cells, distinguished by which cytokines they primarily express upon activation. By expressing different cytokines, these cells enact an inflammatory response specific to the type of pathogen in question. The helper T cells are summarized below:
| Helper T Type | Activating Molecule | Transcriptional Regulator | Cytokines Produced | Function | Defends Against |
|---|---|---|---|---|---|
| Th1 | IL-12 | T-bet | IFN-ɣ IL-2 | Macrophage activation Cytotoxic T lymphocyte activation | Intracellular pathogens |
| Th2 | IL-4 | GATA-3 | IL-4 IL-5 IL-13 | B cell activation for an IgE response Eosinophil activation | Parasites |
| Th17 | IL-6 TGF-β IL-23 | RORɣt | IL-17 IL-22 | Neutrophil activation | Extracellular pathogens |
Cytotoxic T Cells (CD8⁺)
Cytotoxic T cells directly kill infected cells through secretion of perforin and granzyme, similar to an NK cell. CD8 binds to MHCI molecules, which allows these T lymphocytes to bind to any nucleated cell expressing their specific antigen. These antigens are typically proteins associated with intracellular pathogens or neoplasia, allowing for a robust immune response against these types of damage.
Treg Cells
T-regulatory cells are a type of helper T cell, although they do not fit into the traditional classifications above. These cells actually have an immunosuppressive function, and are used in the removal of self-reacting helper T lymphocytes. They are distinguished from their other helper T cousins by expressing CD25. They are discussed further in Immunologic Tolerance.
B Lymphocytes
B lymphocytes mature in the bone marrow, again giving them their “B” name. After the initial maturation, B lymphocytes migrate to the spleen, tonsils and lymph nodes, where they wait for antigen presentation. In these tissues, they are found within lymphoid follicles.
The BCR
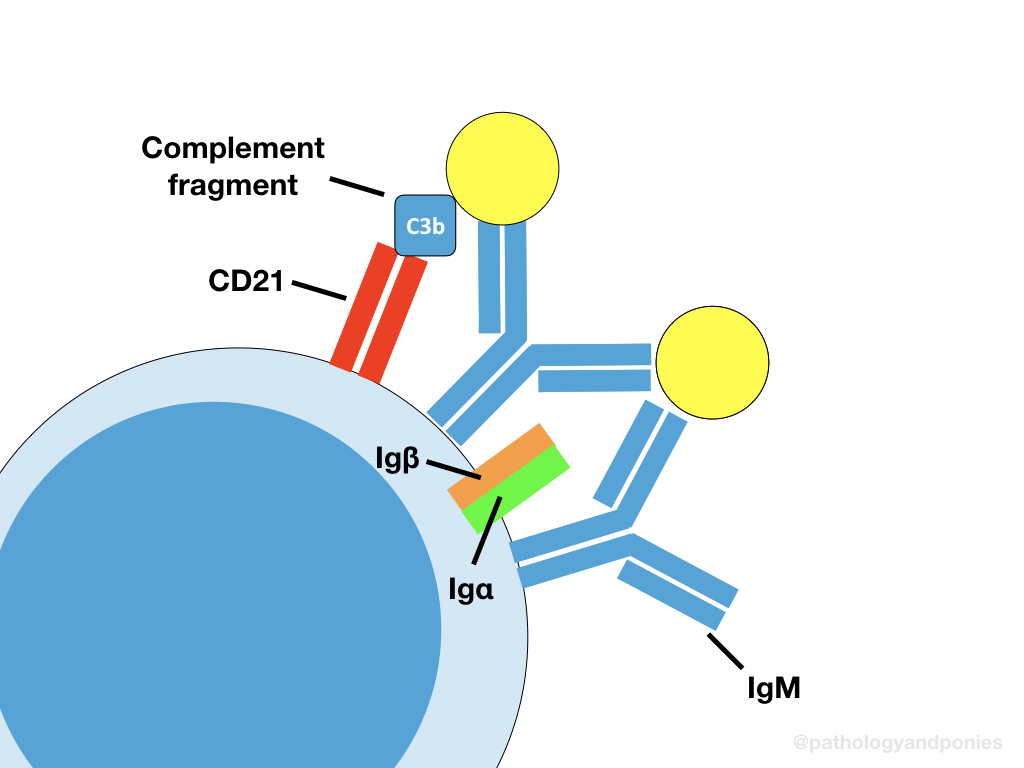
A B-cell receptor is just a membrane-bound immunoglobulin, which binds to its specific antigen and activates the B cell. These bound immunoglobulins are either IgD or IgM. Similar to TCRs, the antigen specificity of immunoglobulins depends on somatic rearrangement. This is further discussed in Immunologic Tolerance.
Each membrane-bound immunoglobulin is associated with accessory proteins called Igα and Igβ, which are invariable. Like the accessory proteins in TCRs, these proteins help with signalling after antigen binding. Unlike most TCRs, the BCR does not require the antigen to be presented to the B cell. This means B cells can bind to soluble antigen, which is important for their function in the spleen.
The BCR also requires a second signal, through binding of CD21. However, because the B cell is not necessarily binding to another cell, the ligand for CD21 are C3 complement fragments, which bind to the soluble antigen. B cells also express CD40 ligand, which binds to CD40 on helper T cells, to receive the signal to switch immunoglobulin type. This is further discussed in Adaptive Immunity Activation.
Tanaka S, Itohara S, Sato M et al. Reduced Formation of Granulomata in γδ T Cell Knockout BALB/c Mice Inoculated with Mycobacterium avium subsp. paratuberculosis. Veterinary Pathology 2000; 37(5):415-421.
Zachary JF. Pathologic Basis of Veterinary Disease, Sixth Edition.
Kumar V, Abbas AK, Aster JC. Robbins and Cotran Pathologic Basis of Disease, Tenth Edition.



